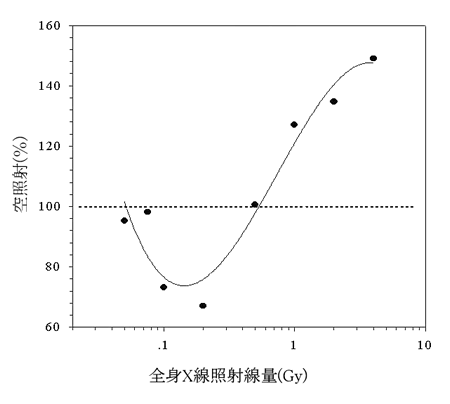
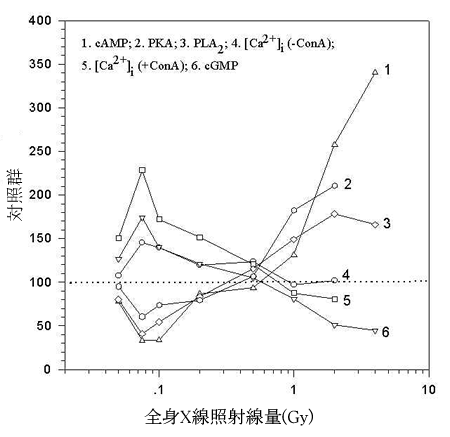
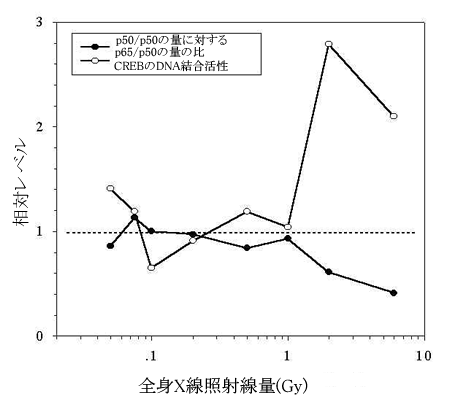
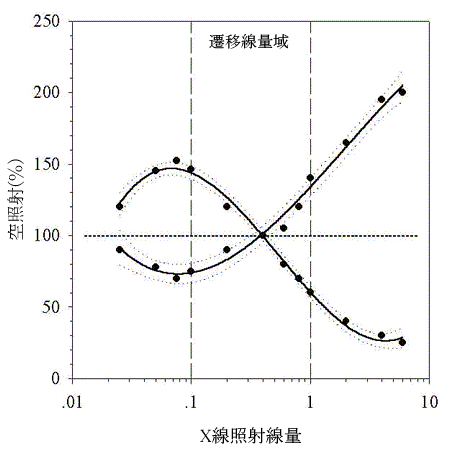
|
1.
|
Liu S-Z. Radiation Hormesis with low-level
exposures. Beijing: Science Publishers, 1996. |
|
2.
|
Li X-Y, Lu Z, Zhang YC, et al. Enhancing
effect of low dose radiation on the proliferative activity of thymocytes
in mice. J Radiat Res Radiat Proc 1994; 12, 124-126. |
|
3.
|
Liu S-Z, Bai O, Chen D, et al. Genes
and protein molecules involved in the cellular activation induced
by low dose radiation. J Radiat Res Radiat Proc 2000; 18: 175-186. |
|
4.
|
Liu S-Z. Biological defense and adaptation
induced by low dose radiation. Human Ecol Risk Assess (HERA) 1998;
4:1217-1254. |
|
5.
|
Liu S-Z. Cellular and molecular basis
of the stimulatory effect of low dose radiation on immunity. In Wei,
L-X, Sugahars, T. and Tao, Z-F. (eds.), High Levels of Natural Radiation.
Amsterdam: Elsevier Science BV, 1997; 341-353. |
|
6.
|
McKenna WG, Maity A, Muschel RJ. Radiation
and the molecular biology of the cell cycle. Radiation Research 1895-1995,
Proc 10th ICRR, Wurzburg, Germany, Aug. 27-Sept. 1, 1995; 629-634. |
|
7.
|
Ye F, Liu S-Z. Effects of whole body
irradiation with X-rays on cell cycle progression of thymocytes in
mice. Radiat Prot 2000; 20:189-192. |
|
8.
|
Liu S-Z, Xie F. Involvement of the Ca2+-protein
kinase C and adenylate cyclase signal pathways in the activation of
thymocytes in response to whole-body irradiation with low dose X-rays.
Chin Med Sci J 2000; 15:1-7. |
|
9.
|
Liu S-Z, Su X, Zhang YC, et al. Signal
transduction in lymphocytes after low dose radiation. Int J Occup
Med Toxicol 1994; 3: 107-117. |
|
10.
|
Danet-Desnoyers G, Meyer MD, Gross TS,
et al. Regulation of endometrial prostaglandin synthesis during early
pregnancy in cattle: effects of phospholipases and calcium in vitro.
Prostaglandins 1995; 50: 313-330. |
|
11.
|
Ghosh S, May MJ, Kopp EB. NF-kappaB
and Rel proteins: evolutionarily conserved mediators of immune response.
Annu Rev Immunol 1998; 16: 225-260. |
|
12.
|
Li X-Y?13. Li X-J?14. Zhang Y?15. et
al. Influence of low dose ionizing radiation on thymic lymphoma induced
by carcinogenic dose radiation in C57BL/6J mice. Acad. Period Abst
Chin 1998; 4: 1406-1407. |
|
16.
|
Li X-J, Yang Y, Li X-Y, et al. Immunologic
mechanism of the suppressive effect of low dose radiation on thymic
lymphoma induced by radiation. J Radiat Res Radiat Proc 1999; 17:
125-128. |